文章導讀
eccDNA是染色體外的一種特殊的環狀DNA,從它的出現至今已長達數年,但在起初的很長一段時間里并未得到人們的重視。隨著高通量技術的發展,eccDNA(extrachromosomal
circular
DNAs,eccDNAs)作為染色體外的環狀DNA的研究也進一步深入。在國際學術期刊《Nature》和《Cell》上相繼發表的關于eccDNA在腫瘤的發生和發展中具有重要功能的報道使eccDNA成為了基因組學研究中的一個爆點。這兩篇文章中認為eccDNA能夠促進腫瘤細胞中原癌基因的表達,這一重要發現使eccDNA成為科研界關注的熱點話題。
然而關于eccDNA在腫瘤細胞內的空間位置及作用機理仍未可知。近期,美國斯坦福大學研究團隊在bioRxiv
預先發表了一篇題目為“EcDNA hubs drive cooperative intermolecular oncogene
expression”的文章,對癌癥細胞中ecDNA
hubs(ecDNA聚集成簇)在致癌基因的表達中發揮重要的作用提出了一個新的分子機制,這為解析ecDNA在癌癥細胞中的作用提出一個新方向,也為癌癥治療提供了新的理論基礎。

發表平臺: bioRxiv
發表日期: 2020.11.20
實驗方法: WGS, RNA-seq, ChIP(以上服務云序均可提供)
文章鏈接:EcDNA hubs drive cooperative intermolecular oncogene expression
研究內容
此前研究發現ecDNA是大小約100kb-幾Mb的雙鏈環狀DNA。由于其自身沒有著絲粒,因此在細胞分裂中可隨機分配到子細胞,這一特性使ecDNA在腫瘤耐藥性方面發揮作用。而且,ecDNA也可以重新整合到染色體上或者形成重復序列(HSRs)。此外,ecDNA相比于染色體DNA有更加松散的結構,更適于其攜帶的致癌基因的轉錄和表達。ecDNA存在于正常染色體之外,其在細胞核中的空間組織目前還不清楚。但值得注意的是,ecDNA在某些生物學過程之后會發生聚集。本文中作者展示了由10-100個ecDNA聚集組成的ecDNA簇,它們聚集在間期細胞核內,驅動分子間增強子的重排從而驅動致癌基因表達。本研究中,通過檢測在MYC-PVT1融合基因的ecDNA空間、表觀和轉錄水平的動態變化,發現ecDNA hubs作為一個組合的增強子集合體,對致癌基因的轉錄發揮調控。
研究結果
(1)ecDNA hubs是癌基因轉錄的主要位點
為了了解ecDNA在轉錄過程中的空間背景,作者使用DNA熒光原位雜交技術(FISH)分別在前列腺癌(PC3)、結直腸癌(COLO320-DM)、膠質母細胞瘤(HK359)以及胃癌(SNU16)中可視化了ecDNA在間期期間在細胞核中的定位,發現這些ecDNA是幾十到幾百個聚集在間期細胞的細胞核的特定區域,這表明ecDNA
相互之間具有很強的聚集性,稱之為ecDNA hubs。為了評估ecDNA
hubs的轉錄活性,作者結合了DNA和新生RNA利用FISH檢測了在PC3和COLO320-DM細胞系中MYC等位基因的轉錄活性,發現ecDNA
hubs上致癌基因的轉錄的概率明顯高于染色體位點,且發現MYC轉錄概率與ecDNA聚類之間有顯著的相關性。因此,當更多的ecDNA在同一細胞聚集時,ecDNA
hubs更有可能轉錄癌基因。

(2)單細胞水平識別與強效癌基因表達相關的ecDNA增強子
為了了解ecDNA上致癌基因表達的調控,作者在單細胞水平上鑒定了與癌基因高表達相關的ecDNA上的調控元件并檢測了癌細胞的異質性,以及ecDNA與癌基因轉錄的關系。作者采用單細胞組學的方法,使用ATAC-
seq和RNA-seq共檢測到結直腸癌細胞系col320
-DM和COLO320-HSR中72049個細胞獲得的轉錄組和染色質開放性圖譜。進一步整合了每個細胞的轉錄組和染色質開放性圖譜,發現MYC的開放性隨RNA表達而增加。MYC在COLO320-DM中的表達與開放性相比于COLO320-HSR是高度異質的。這些結果表明,調控元件可以影響ecDNA在癌基因表達中的細胞間差異。為了鑒定細胞中表達高水平MYC的ecDNA上的活性調控元件,ATAC-seq比較表明,高MYC表達的COLO320-DM細胞既包含更高的ecDNA拷貝數,也包含更多的調控元件,在ecDNA上的差異峰分析鑒定出47個與MYC高表達密切相關的調控元件,而在染色體HSRs上鑒定出5個調控元件,這些結果表明,MYC
轉錄的增加與ecDNA中許多調節元件有關。總之,這些結果表明,細胞間增強子的差異可能是ecDNA 癌基因表達異質性的關鍵。
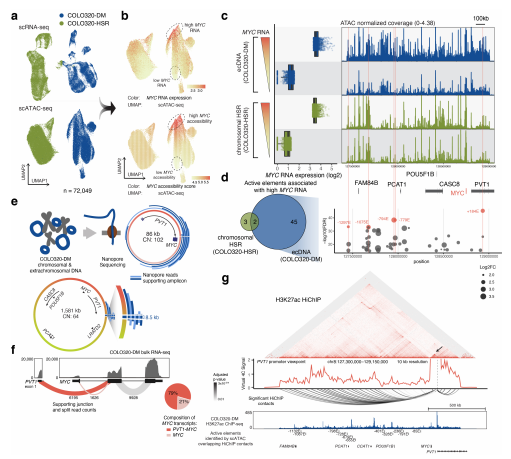
(3)增強子和啟動子之間的分子間互作促進ecDNA 的異質化和致癌基因的表達
由于發現(i)細胞核內的ecDNA hubs與活躍的癌基因表達相關,(ii)
ecDNA的癌基因表達與差異調節元件相關,作者下一步研究這些差異元件是否參與不同的ecDNA分子之間的增強子-啟動子相互作用。作者整合了WGS、單分子DNA測序和3D
增強子連接體圖譜,分析ecDNA
hubs對致癌基因表達的調控。先根據之前的WGS的數據,檢測COLO320-DM細胞中ecDNA在MYC位點有重排,確定了PVT1能和MYC的外顯子2和3發生融合,并采用nanopore三代單分子測序的方法進行了驗證,結果顯示ecDNA分子在COLO320-DM中存在高度的異質性。隨后采用HiChIP檢測了COLO320-DM和COLO320-HSR中H3K27ac與激活型增強子和ecDNA的作用,發現在ecDNA的拷貝數高的COLO320-DM中H3K27ac與激活型增強子結合越緊密。此外,通過ATAC-seq鑒定了大量的激活型增強子,結果發現其與MYC基因有相互作用。綜上結果,作者認為增強子和啟動子之間的互作促進ecDNA的異質化和致癌基因的表達。

(4)BRD4連接ecDNA hubs并驅動癌基因轉錄
廣泛的遠距離接觸和H3K27ac相關的DNA接觸提高了BET蛋白可能參與ecDNA
hubs轉錄的可能性。BET是染色質閱讀蛋白,為了檢測BET蛋白在ecDNA促進轉錄作用,檢測在兩種細胞中MYC位點BRD4的定位情況。根據H2K27ac,BRD4的ChIP-seq結果和ATAC-seq數據,表明H3K27ac
存在的激活的ecDNA增強子,與BRD4同時存在,BRD4連接ecDNA hubs,促進致癌基因的轉錄。為了評估BET抑制劑JQ1在ecDNA
hubs的功能中作用,作者進行了JQ1處理,發現加入JQ1后ecDNA出現分散,且具有細胞特異性。而加入RNA
PolII抑制劑,降低MYC的轉錄但不影響ecDNA hubs的聚集。通過新生RNA和DNA
FISH和BRD4的ChIP-seq結果表明ecDNA hubs的形成依賴于BET蛋白,而ecDNA
hubs的破壞會引起MYC致癌基因轉錄的抑制和ecDNA癌細胞的死亡。除了BET的抑制劑會抑制ecDNA的轉錄,作者還發現CRISPR的干擾劑,也可以抑制PVT1的啟動子,降低PVT1-MYC的轉錄從而降低總的MYC的RNA水平。這一結果顯示ecDNA可能是調控致癌基因表達的一個更有效的思路。
